1.3 ОБРАБОТКА ЗРИТЕЛЬНОГО СИГНАЛА
Процесс нейронной обработки зрительной информации на уровне сетчатки сложен, а на дальнейших стадиях он становится значительно (если не бесконечно) более сложным. Данный раздел - это краткий обзор путей, по которым идет зрительная информация, и имеет смысл начать его с общей карты этих путей.
Оптическое изображение на сетчатке вначале преобразуется в химические и электрические сигналы фоторецепторов, а затем эти сигналы обрабатываются клетками нейронной сети сетчатки (горизонтальными, биполярными, амак-риновыми и ганглиозными клетками, описанными выше). Аксоны ганглиозных клеток формируют зрительный нерв, который идет к т.н. латеральному
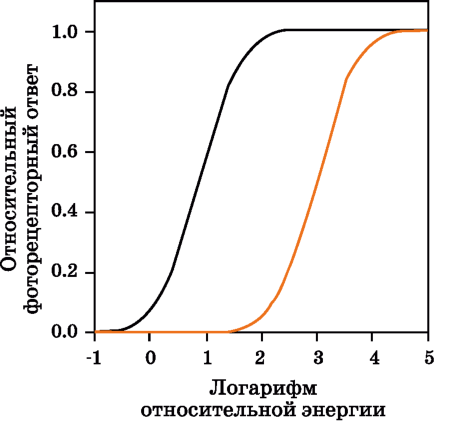
Рис. 1.8 Относительные энергетические ответы палочковых и колбочковых фоторецепторов.
коленчатому телу (lateral geniculate nucleus - LGN) в таламусе1. Клетки LGN, получив входной сигнал от ганглиозных клеток, отсылают его в т.н. первую зрительную область (V1) затылочных долей коры головного мозга, в которых обработка информации становится невероятно сложной. В коре головного мозга было найдено почти 30 зрительных областей, названные затем как V2, V3, V4, MT и т.д. Сигналы от этих областей отсылаются к некоторым другим областям и обратно. Корковая обработка включает в себя множество стадий: прямую, обратную и латеральную обработки; где-то в недрах этой сети и формируется наше окончательное восприятие. Кое-какие детали этих процессов описаны в следующих параграфах.
Свет, воздействующий на сетчатку, абсорбируется фотопигментом различных фоторецепторов (в палочках, в частности, это родопсин). После поглощения фотона родопсин меняет свою структуру, происходит цепная химическая реакция, итогом которой является закрытие ионных каналов в клеточной стенке, продуцирующее электрический сигнал, основанный на разности относительных концентраций различных ионов (к примеру, натриевых и карбонатных) вне и внутри клетки. Аналогичные процессы протекают в колбочках, родопсин которых состоит из опсина и ретиналя: колбочки всех трех типов имеют сходную фотопигментную структуру, но у колбочковых опсинов («конопси-нов») немного разные молекулярные формулы, что ведет к отличиям в спектральной чувствительности колбочек. Каждый тип колбочек (L, M или £) содержит свою форму «конопсина».
На рис. 1.8 показаны относительные ответы фоторецепторов как функция световой экспозиции сетчатки. Любопытно, что характеристики этих функций подобны характеристикам многих устройств ввода и вывода изображений.
В нижней части графика виден порог, ниже которого рецепторы не отвечают. Затем следует более или менее линейный участок кривых, и за ним - финальный участок - зона насыщения (верхний конец кривых). Приведенные кривые описывают рецепторный фототок и демонстрируют самую первую стадию зрительной обработки. В дальнейшем сигналы обрабатываются нейронами сетчатки и синапсами. Результатом этой обработки является преобразованный сигнал, который ганглиозные клетки отправляют по зрительному нерву в мозг.
Рецептивные поля
По ряду причин (таких, как необходимость подавления шумов и обеспечение высокой скорости передачи) амплитудно-модулированные сигналы фоторецепторов конвертируются ганглиозными клетками и вышестоящими отделами мозга в частотно-модулированное представление. Сегодня несомненно то, что величина сигнала от возбужденной нервной клетки представлена количеством пиков напряжения в секунду, а не разностью потенциалов по обе стороны клеточной стенки.