«_» «_» О U U U т~*
ответа на светлый-темный, красный-зеленый и желтый-синий. В то время такое утверждение казалось физиологически абсурдным, и оппонентная теория Геринга не получила признания.
Современная теория оппонентных цветов
В середине XX-го века оппонентная теория возродилась, поскольку стали появляться подтверждающие ее экспериментальные данные, к примеру: Свэтичин (1956) обнаружил оппонентные сигналы при электрофизиологических измерениях ответов в сетчатке золотой рыбки (которые оказались трихроматически-ми!), Де'Валойс с коллегами (1958) обнаружил подобные оппонентные физиологические ответы в LGN-клетках макаки, Джеймсон и Гурвич (1955) представили количественные психофизические данные, полученные на основе т.н. безотте-ночных экспериментов с людьми. Эксперименты Джеймсона и Гурвича позволили измерить относительные спектральные чувствительности оппонентных путей. Все эти данные, объединенные с огромным количеством дополнительных исследований, привели к развитию современной оппонентной теории цветового зрения, которую иногда называют стадийной теорией (см. рис. 1.13).
Рис. 1.13 демонстрирует, что на первой стадии цветового зрения рецепторы, как и предполагали Максвелл, Юнг и Гельмгольц, действительно трихромати-ческие. Однако вопреки простой трихроматической теории три «цветоделен-ных» изображения не передаются непосредственно в мозг: вместо этого нейроны сетчатки (и возможно, более высокие отделы зрительной системы) кодируют цвет через оппонентные сигналы. Выходные сигналы от всех трех типов колбочек суммируются (L+M+S) с целью получения ахроматического ответа, который до тех пор соответствует кривой CIE V(X), пока суммация берется в пропорции к относительной заселенности сетчатки тремя типами колбочек. Различия в колбочковых сигналах позволяют сконструировать краснозеленые (L-M+S) и желто-синие (L+M-S) оппонентные сигналы.
Преобразование LMS-сигналов в оппонентные приводит к декорреляции цветовой информации, идущей по трем каналам, тем самым обеспечивая более эффективную передачу сигналов и снижая шумовые помехи. Три оппонентных пути, разумеется, имеют индивидуальные пространственные и временные характеристики, очень важные для прогнозирования цветового восприятия (но их мы обсудим в разделе 1.5).
Значимость преобразования трихроматических сигналов в оппонентные отражена в формулировках всех моделей цветового восприятия. В рис. 1.13 включена не только схематическая диаграмма нейронного «телеграфа», который продуцирует оппонентные ответы, но также и относительные спектральные чувствительности оппонентных механизмов до и после оппонентного кодирования.
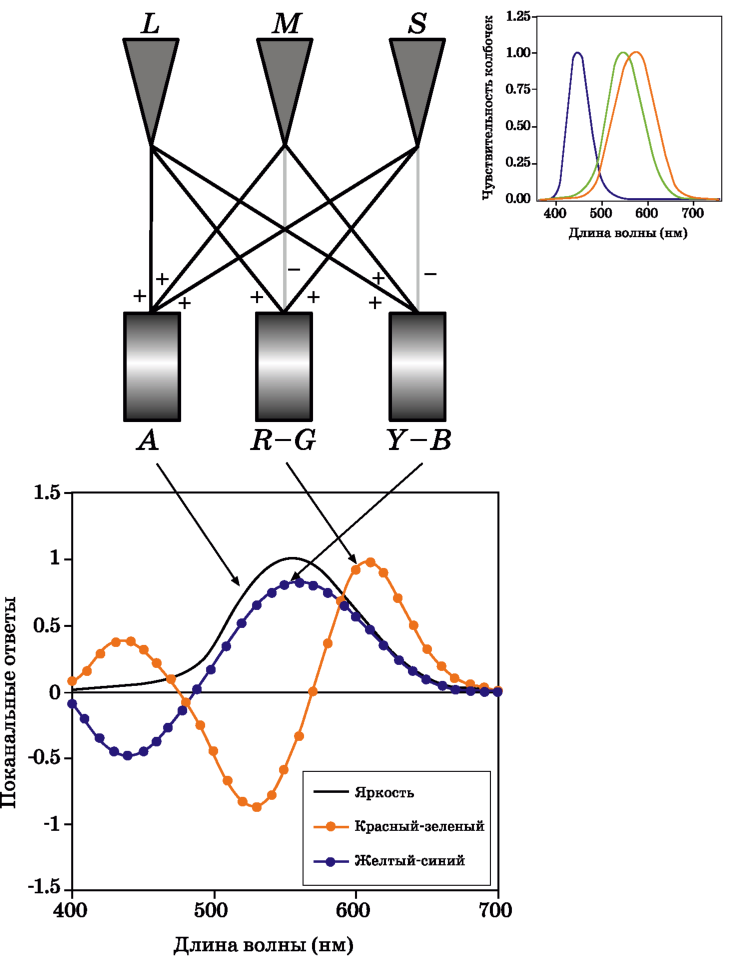
Рис. 1.13 Схематическая иллюстрация кодирования колбочковых сигналов в оппонентные цветовые сигналы в зрительной системе человека.
Механизмы адаптации
Заметим, что было бы неправомерно рассматривать обработку цветовых сигналов человеческой зрительной системой лишь как статическую: необходимо еще разобрать динамические приспособительные механизмы, которые служат оптимизации визуального ответа в тех или иных условиях просмотра, то есть изучить различные виды адаптации.